

Modified Chaihu Longgu Muli Decoction Improved the Synaptic Plasticity of PFC and Alleviated the Depressive Symptoms in CKD Mice via p38 MAPK Pathway
-
摘要: 目的 探讨柴胡加龙骨牡蛎汤加减(CLMD)对慢性肾脏病(CKD)小鼠抑郁行为的作用及可能机制。方法 3月龄雄性C57BL/6小鼠60只, 分为对照组、模型组、氟西汀组、CLMD低剂量组、CLMD中剂量组和CLMD高剂量组。除对照组外, 其余5组小鼠先予0.2%腺嘌呤饮食喂养2周。中药干预组分别予2.36、4.725、9.45 g·kg-1·d-1的CLMD灌胃, 氟西汀组予10 mg·kg-1·d-1氟西汀灌胃, 对照组、模型组予生理盐水灌胃, 共计6周。糖水偏好测试、强迫游泳实验检测小鼠抑郁行为表型, Western blot法检测前额叶皮层(PFC) p38丝裂原活化蛋白激酶(p38 MAPK)、磷酸化p38 MAPK(p-p38 MAPK)、脑源性神经营养因子(BDNF)、环磷腺苷效应元件结合蛋白(CREB)、磷酸化CREB(p-CREB)、突触后致密蛋白95(PSD95)、突触素(SYN)、生长相关蛋白43(GAP43)和突触囊泡膜蛋白(SYP)表达; 免疫荧光法检测PFC脑区核内p-CREB的表达; Golgi-Cox染色检测PFC树突棘密度; ELISA法检测外周及PFC炎性因子水平及血清肌酐、尿素氮水平。结果 行为学检测结果显示: 与对照组相比, 模型组不动时间增加,糖水偏好指数下降(P < 0.01);与模型组相比, 给予中、高剂量CLMD干预后小鼠不动时间下降(P < 0.01), 糖水偏好指数增加(P < 0.05, P < 0.01), CLMD中、高剂量组与氟西汀组之间差异无统计学意义(P>0.05)。Western blot结果显示: 与对照组相比, 模型组小鼠PFC中p-p38 MAPK/p38 MAPK表达增加(P < 0.01), BDNF、p-CREB/CREB、PSD95、SYN、GAP43、SYP表达下调(P < 0.01);与模型组相比, CLMD中、高剂量组, 氟西汀组小鼠p-p38 MAPK/p38 MAPK表达下调(P < 0.01), BDNF、p-CREB/CREB、PSD95、SYN、GAP43、SYP表达增加(P < 0.01)。免疫荧光结果显示: 与对照组相比, 模型组PFC的p-CREB核内荧光水平明显降低(P < 0.01);与模型组相比, CLMD中、高剂量组, 氟西汀组p-CREB核内荧光强度增加(P < 0.05, P < 0.01)。Golgi-Cox染色结果显示: 与对照组相比, 模型组小鼠PFC树突棘密度降低(P < 0.01);与模型组相比, CLMD中、高剂量组, 氟西汀组小鼠PFC树突棘密度增加(P < 0.01)。ELISA法检测结果显示: 与对照组相比, 模型组PFC及外周血清中炎性因子水平增加(P < 0.01), 血清尿素氮、肌酐水平增加(P < 0.01);与模型组相比, 氟西汀组及CLMD中、高剂量组PFC及外周血清中炎性因子水平下降(P < 0.05, P < 0.01), CLMD中、高剂量组血清尿素氮、肌酐水平下降(P < 0.05, P < 0.01), 氟西汀组尿素氮、肌酐水平无明显变化。结论 CLMD能调节中枢炎症状态, 改善PFC突触可塑性, 减轻CKD小鼠抑郁症状,同时可能通过影响外周炎症状态延缓CKD小鼠肾脏功能恶化。Abstract: OBJECTIVE To study the effect and mechanism of modified Chaihu Longgu Muli Decoction (CLMD) on depressive behavior in chronic kidney disease (CKD) mice.METHODS Sixty 3-month-old male C57BL/6 mice were divided into control group, model group, fluoxetine group, CLMD low dose group, CLMD medium dose group and CLMD high dose group. Except the control group, the rest 5 groups of mice were intervened by 0.2% adenine diet for 2 weeks, then CLMD low, medium, high groups were gavaged 2.36, 4.725, 9.45 g·kg-1·d-1 of CLMD, respectively. Fluoxetine group was given 10 mg·kg-1·d-1 fluoxetine by gavage, while the control group and model group were given the same amount of normal saline, for six weeks. Sugar preference test and forced swimming test were used to evaluate the depressive state of mice, Western blot was used to detect the expressions of p38 mitogen-activated protein kinase (p38 MAPK), phosphorylated p38 MAPK (p-p38 MAPK), brain derived neurotrophic factor (BDNF), cAMP-response element binding protein (CREB), phosphorylation CREB (p-CREB), postsynaptic density protein 95 (PSD95), Synapsin (SYN) and growth associated protein 43 (GAP43), Synaptophysin (SYP) proteins in the prefrontal cortex (PFC). The nucleus expression of p-CREB in PFC was detected by immunofluorescence assay, the changes of dendrite spines in PFC were detected by Golgi-Cox staining, and the levels of inflammatory factors in peripheral serum and PFC, and the serum creatinine and urea nitrogen were detected by ELISA assay.RESULTS The behavior test results showed that, compared with the control group, the immobility time of the model group increased and the percentage of sugar preference level decreased (P < 0.01). Compared with the model group, the immobility time of mice decreased and the percentage of sugar water preference increased after the intervention of CLMD (P < 0.05, P < 0.01), there was no significant difference among the medium dose group, the high dose group and fluoxetine group (P>0.05). Western blot results showed that, compared with the control group, the expressions of p-p38 MAPK/p38 MAPK in PFC of the model group increased (P < 0.01), and the expressions of BDNF, p-CREB/CREB, PSD95, SYN, GAP43 and SYP down-regulated (P < 0.01). Compared with the model group, the relative expressions of p-p38 MAPK/p38 MAPK in medium-dose, high-dose groups and fluoxetine group down-regulated (P < 0.01), and the relative expressions of BDNF, p-CREB/CREB, PSD95, SYN, GAP43 and SYP increased (P < 0.01). Immunofluorescence results showed that, compared with the control group, the nuclear fluorescence level of p-CREB in PFC of the model group significantly decreased (P < 0.01). Compared with the model group, the nuclear fluorescence expression of p-CREB increased in medium-dose, high-dose groups and fluoxetine group (P < 0.05, P < 0.01). The results of Golgi-Cox staining showed that the number of PFC dendritic spines in model group was lower than that in control group (P < 0.01). Compared with the model group, the numbers of PFC dendritic spines increased after medium-dose, high-dose CLMD and fluoxetine intervention (P < 0.01). ELISA results showed that, compared with the control group, the levels of inflammatory factors in both peripheral blood and PFC of model group increased (P < 0.01), the levels of serum urea nitrogen and creatinine increased (P < 0.01). Compared with model group, after medium-dose, high-dose CLMD intervention, the levels of pro-inflammatory factors, serum urea nitrogen and creatinine in PFC and peripheral blood decreased (P < 0.05, P < 0.01).CONCLUSION CLMD can alleviate the depressive symptoms of CKD mice by regulating the inflammatory state of the central nervous system and affecting the synaptic plasticity of PFC. Meanwhile, CLMD can delay the deterioration of kidney function in CKD model mice by regulating the inflammatory state of the peripheral.
-

图 1 CLMD对CKD小鼠抑郁状态的影响
注: 与对照组比较, ##P < 0.01;与模型组比较, *P < 0.05, **P < 0.01;与氟西汀组相比, & P < 0.05。x±s, n=10。

图 2 CLMD对CKD小鼠PFC炎症状态的影响
注: 与对照组比较, ##P < 0.01;与模型组比较, *P < 0.05, **P < 0.01。x±s, n=6。

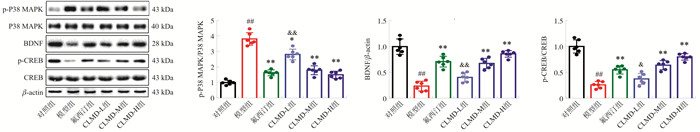
图 3 各组小鼠PFC内p-p38 MAPK/p38 MAPK、BDNF、p-CREB/CREB蛋白相对表达水平比较
注: 与对照组比较, ##P < 0.01;与模型组比较, *P < 0.05, **P < 0.01;与氟西汀组相比, & P < 0.05, $ & P < 0.01。x±s, n=6。

图 4 各组小鼠PFC脑区p-CREB免疫荧光染色图
注: 与对照组比较, ##P < 0.01;与模型组比较, *P < 0.05, **P < 0.01;与氟西汀组相比, & & P < 0.01。x±s, n=4。

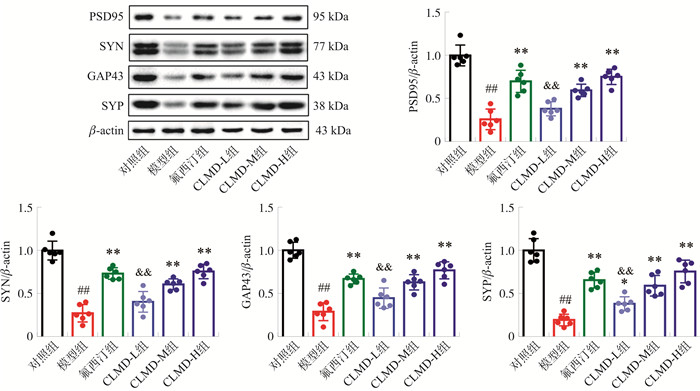
图 5 各组小鼠PFC突触相关蛋白相对表达水平的比较
注: 与对照组比较, ##P < 0.01;与模型组比较, *P < 0.05, **P < 0.01;与氟西汀组相比, & & P < 0.01。x±s, n=6。

图 6 各组小鼠PFC脑区Golgi-Cox染色显示神经元树突棘密度
注: 与对照组比较, ##P < 0.01;与模型组比较, **P < 0.01;与氟西汀组相比, & & P < 0.01。x±s, n=3。

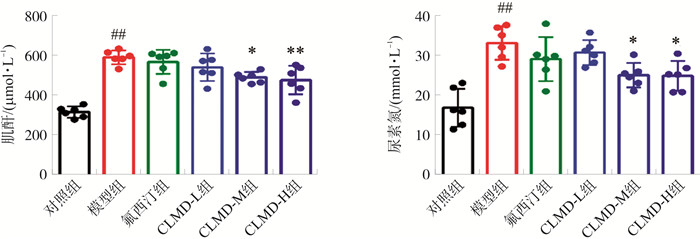
图 7 CLMD对CKD小鼠外周炎性因子水平的影响
注: 与对照组比较, ##P < 0.01;与模型组比较, *P < 0.05, **P < 0.01。x±s, n=6。
-
[1] ORTIZ A, COVIC A, FLISER D, et al. Epidemiology, contributors to, and clinical trials of mortality risk in chronic kidney failure[J]. Lancet, 2014, 383(9931): 1831-1843. doi: 10.1016/S0140-6736(14)60384-6 [2] JHA V, GARCIA-GARCIA G, ISEKI K, et al. Chronic kidney disease: Global dimension and perspectives[J]. Lancet, 2013, 382(9888): 260-272. doi: 10.1016/S0140-6736(13)60687-X [3] SUN L, XU H, WANG Y, et al. The mitochondrial-targeted peptide SBT-20 ameliorates inflammation and oxidative stress in chronic renal failure[J]. Aging, 2020, 12(18): 18238-18250. doi: 10.18632/aging.103681 [4] PALMER S, VECCHIO M, CRAIG JC, et al. Prevalence of depression in chronic kidney disease: Systematic review and meta-analysis of observational studies[J]. Kidney Int, 2013, 84(1): 179-191. doi: 10.1038/ki.2013.77 [5] SIMÕES E SILVA AC, MIRANDA AS, ROCHA NP, et al. Neuropsychiatric disorders in chronic kidney disease[J]. Front Pharmacol, 2019, 10: 932. doi: 10.3389/fphar.2019.00932 [6] CHIANG HH, GUO HR, LIVNEH H, et al. Increased risk of progression to dialysis or death in CKD patients with depressive symptoms: A prospective 3-year follow-up cohort study[J]. J Psychosom Res, 2015, 79(3): 228-232. doi: 10.1016/j.jpsychores.2015.01.009 [7] WALTHER CP, SHAH AA, WINKELMAYER WC. Treating depression in patients with advanced CKD: Beyond the generalizability frontier[J]. JAMA, 2017, 318(19): 1873-1874. doi: 10.1001/jama.2017.17553 [8] 刘渡舟. 伤寒论校注[M]. 北京: 人民卫生出版社, 2013: 85-86. [9] LI LF, LU J, LI XM, et al. Antidepressant-like effects of the saponins extracted from Chaihu-Jia-Longgu-Muli-Tang in a rat unpredictable chronic mild stress model[J]. Fitoterapia, 2012, 83(1): 93-103. doi: 10.1016/j.fitote.2011.09.017 [10] WANG X, CHEN J, ZHANG HL, et al. Immediate and persistent antidepressant-like effects of Chaihu-Jia-Longgu-Muli-Tang are associated with instantly up-regulated BDNF in the hippocampus of mice[J]. Biosci Rep, 2019, 39(1): BSR20181539. doi: 10.1042/BSR20181539 [11] 芮轶群, 吕阳, 李文婷, 等. 柴胡加龙骨牡蛎汤加减治疗失眠的meta分析[J]. 山东中医杂志, 2019, 38(12): 1123-1130, 1136. https://www.cnki.com.cn/Article/CJFDTOTAL-SDZY201912013.htm [12] SANTANA AC, DEGASPARI S, CATANOZI S, et al. Thalidomide suppresses inflammation in adenine-induced CKD with uraemia in mice[J]. Nephrol Dial Transplant, 2013, 28(5): 1140-1149. doi: 10.1093/ndt/gfs569 [13] PULSKENS WP, VERKAIK M, SHEEDFAR F, et al. Deregulated renal calcium and phosphate transport during experimental kidney failure[J]. PLoS ONE, 2015, 10(11): e0142510. doi: 10.1371/journal.pone.0142510 [14] MOTOHASHI H, TAHARA Y, WHITTAKER DS, et al. The circadian clock is disrupted in mice with adenine-induced tubulointerstitial nephropathy[J]. Kidney Int, 2020, 97(4): 728-740. doi: 10.1016/j.kint.2019.09.032 [15] KATON WJ. Epidemiology and treatment of depression in patients with chronic medical illness[J]. Dialogues Clin Neurosci, 2011, 13(1): 7-23. doi: 10.31887/DCNS.2011.13.1/wkaton [16] IWAGAMI M, TOMLINSON LA, MANSFIELD KE, et al. Prevalence, incidence, indication, and choice of antidepressants in patients with and without chronic kidney disease: A matched cohort study in UK Clinical Practice Research Datalink[J]. Pharmacoepidemiol Drug Saf, 2017, 26(7): 792-801. doi: 10.1002/pds.4212 [17] DOOLEY LN, KUHLMAN KR, ROBLES TF, et al. The role of inflammation in core features of depression: Insights from paradigms using exogenously-induced inflammation[J]. Neurosci Biobehav Rev, 2018, 94: 219-237. doi: 10.1016/j.neubiorev.2018.09.006 [18] KÖHLER CA, FREITAS TH, MAES M, et al. Peripheral cytokine and chemokine alterations in depression: A meta-analysis of 82 studies[J]. Acta Psychiatr Scand, 2017, 135(5): 373-387. doi: 10.1111/acps.12698 [19] MILLER AH. Beyond depression: The expanding role of inflammation in psychiatric disorders[J]. World Psychiatry, 2020, 19(1): 108-109. doi: 10.1002/wps.20723 [20] JING W, JABBARI B, VAZIRI ND. Uremia induces upregulation of cerebral tissue oxidative/inflammatory cascade, down-regulation of Nrf2 pathway and disruption of blood brain barrier[J]. Am J Transl Res, 2018, 10(7): 2137-2147. https://europepmc.org/article/MED/30093950 [21] MAZUMDER MK, GIRI A, KUMAR S, et al. A highly reproducible mice model of chronic kidney disease: Evidences of behavioural abnormalities and blood-brain barrier disruption[J]. Life Sci, 2016, 161: 27-36. doi: 10.1016/j.lfs.2016.07.020 [22] PANDEY GN, RIZAVI HS, BHAUMIK R, et al. Chemokines gene expression in the prefrontal cortex of depressed suicide victims and normal control subjects[J]. Brain Behav Immun, 2021, 94: 266-273. doi: 10.1016/j.bbi.2021.01.033 [23] WANG H, HUANG B, WANG W, et al. High urea induces depression and LTP impairment through mTOR signalling suppression caused by carbamylation[J]. EBioMedicine, 2019, 48: 478-490. doi: 10.1016/j.ebiom.2019.09.049 [24] DUMAN RS, AGHAJANIAN GK, SANACORA G, et al. Synaptic plasticity and depression: New insights from stress and rapid-acting antidepressants[J]. Nat Med, 2016, 22(3): 238-249. doi: 10.1038/nm.4050 [25] LI MX, ZHENG HL, LUO Y, et al. Gene deficiency and pharmacological inhibition of caspase-1 confers resilience to chronic social defeat stress via regulating the stability of surface AMPARs[J]. Mol Psychiatry, 2018, 23(3): 556-568. doi: 10.1038/mp.2017.76 [26] TONG L, PRIETO GA, KRAMÁR EA, et al. Brain-derived neurotrophic factor-dependent synaptic plasticity is suppressed by interleukin-1β via p38 mitogen-activated protein kinase[J]. J Neurosci, 2012, 32(49): 17714-17724. doi: 10.1523/JNEUROSCI.1253-12.2012 [27] ZHANG X, LEI B, YUAN Y, et al. Brain control of humoral immune responses amenable to behavioural modulation[J]. Nature, 2020, 581(7807): 204-208. doi: 10.1038/s41586-020-2235-7 [28] FAN KQ, LI YY, WANG HL, et al. Stress-induced metabolic disorder in peripheral CD4+T cells leads to anxiety-like behavior[J]. Cell, 2019, 179(4): 864-879. doi: 10.1016/j.cell.2019.10.001 -
 下载:
下载:




 点击查看大图
点击查看大图

计量
- 文章访问数: 185
- HTML全文浏览量: 51
- PDF下载量: 114
- 被引次数: 0
