

Baitouweng Decoction Alleviates DSSInduced Colitis via Rebalancing Gut Microbiota and Regulating Short Chain Fatty Acids
-
摘要: 目的 探究白头翁汤对溃疡性结肠炎(Ulcerative colitis, UC)模型小鼠的治疗作用及可能机制。方法 采用葡聚糖硫酸钠(Dextran sulphate sodium, DSS)诱导UC模型小鼠, 给予白头翁汤干预。实验过程中, 每日记录小鼠体质量、腹泻、便血情况。实验结束后测量小鼠结肠长度, HE染色观察结肠病理变化, qPCR检测结肠炎症因子表达, 16S rRNA检测小鼠肠道菌群组成, GC-MS法检测回盲瓣内容物短链脂肪酸(SCFAs)含量。结果 与正常组小鼠相比, 模型组小鼠体质量下降, 结肠缩短(P < 0.01), 结肠组织中炎性细胞浸润, 结构紊乱, 部分形态消失; 结肠组织中促炎因子IL-1β、IL-6和TNF-α表达增加(P < 0.05, P < 0.01)。肠道菌群结构失衡, 厚壁菌门与拟杆菌门比值降低, 回盲瓣内容物中丁酸、戊酸含量降低(P < 0.01)。与模型组小鼠相比, 白头翁汤干预后, 小鼠体质量明显增加, 结肠长度增加(P < 0.05, P < 0.01), 结肠存在炎性细胞浸润, 但结构清晰, IL-1β、IL-6和TNF-α表达下降(P < 0.05, P < 0.01), 厚壁菌门与拟杆菌门比值增加, 回盲瓣中丁酸、戊酸含量增加(P < 0.05, P < 0.01)。结论 白头翁汤可能通过调节肠道菌群, 改善SCFAs代谢起到治疗溃疡性结肠炎的作用。Abstract: OBJECTIVE To explore the treatment and mechanism of Baitouweng decoction on ulcerative colitis mice.METHODS Colitis mice were induced by DSS and treated with or without Baitouweng decoction. During experiment, body weight of mice, the symptom of diarrhea and bloody stool were recorded. At the end of experiment, the length of colon was measured, colonic pathology was observed after HE staining, the expression of cytokines was evaluated by qPCR, the construction of gut microbiota was detected by 16S rRNA, the concentration of short chain fatty acids was measured by gas chromatography-mass spectrometry.RESULTS Compared with the healthy mice, the body weight and the length of colon decreased in colitis mice(P < 0.01), the construction of colon was disorder with infiltration of inflammation cells, and the expressions of IL-1β, IL-6 and TNF-α was upregulated(P < 0.05, P < 0.01). But the expression of anti-inflammatory cytokines were not different between these two groups. Moreover, dysbacteriosis and low level of short chain fatty acids were observed. After Baitouweng decoction treatment, the weight of mice and the length of colon were increased(P < 0.05, P < 0.01). The expressions of pro-inflammatory cytokine, such as IL-1β, IL-6 and TNF-α, was downregulated(P < 0.05, P < 0.01). And the construction of gut microbiota was rebalanced especially in DSS+BTW-H group. The contents of butyric acid and valeric acid increased significantly(P < 0.05, P < 0.01).CONCLUSION Baitouweng decoction has therapeutic effect on DSS-induced colitis through rebalancing gut microbiota and short chain fatty acids.
-
Key words:
- Baitouweng decoction /
- ulcerative colitis /
- gut microbiota /
- short chain fatty acids
-

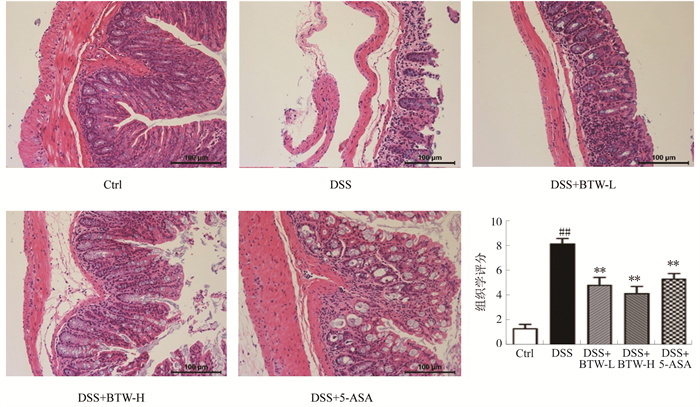
图 1 白头翁汤对DSS诱导模型小鼠体质量、DAI及结肠长度的影响
注: Ctrl.正常组; DSS.模型组; DSS+BTW-L.白头翁汤低剂量组; DSS+BTW-H.白头翁汤高剂量组; DSS+5-ASA.5-ASA组。与Ctrl组相比, #P < 0.05, ##P < 0.01;与DSS组相比, *P < 0.05, * *P < 0.01。

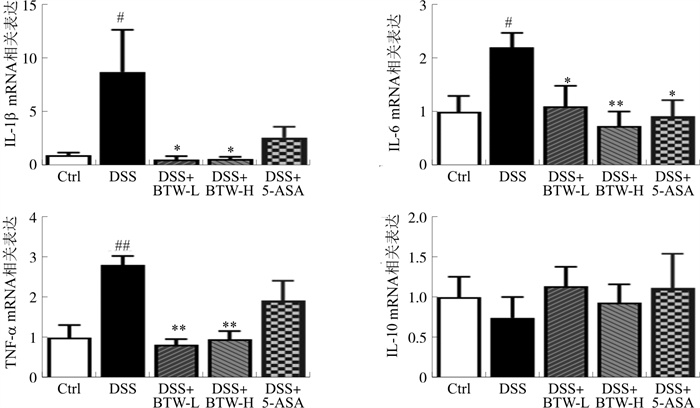
图 3 白头翁汤对DSS诱导肠炎小鼠结肠组织炎症因子表达的影响
注: 与Ctrl组相比, #P < 0.05,##P < 0.01;与DSS组相比, *P < 0.05, * *P < 0.01。x±s,n=6~8。

图 4 白头翁汤对DSS诱导肠炎小鼠肠道菌群结构影响
注: 与Ctrl组相比, ##P < 0.01;与DSS组相比, *P < 0.05, * *P < 0.01。x±s, n=6~8。

图 5 白头翁汤对产SCFAs菌影响
注: 与Ctrl组相比, #P < 0.05, ##P < 0.01;与DSS组相比, *P < 0.05, * *P < 0.01。x±s, n=6~8。
-
[1] COOPER HS, MURTHY SN, SHAH RS, et al. Clinicopathologic study of dextran sulfate sodium experimental murine colitis[J]. Lab Invest, 1993, 69(2): 238-249. http://www.ncbi.nlm.nih.gov/pubmed/8350599 [2] LOUIS P, HOLD GL, FLINT HJ. The gut microbiota, bacterial metabolites and colorectal cancer[J]. Nat Rev Microbiol, 2014, 12(10): 661-672. doi: 10.1038/nrmicro3344 [3] SCOTT KP, MARTIN JC, CAMPBELL G, et al. Whole-genome transcription profiling reveals genes up-regulated by growth on fucose in the human gut bacterium "Roseburia inulinivorans"[J]. J Bacteriol, 2006, 188(12): 4340-4349. doi: 10.1128/JB.00137-06 [4] KOH A, DE VADDER F, KOVATCHEVA-DATCHARY P, et al. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites[J]. Cell, 2016, 165(6): 1332-1345. doi: 10.1016/j.cell.2016.05.041 [5] FURUSAWA Y, OBATA Y, FUKUDA S, et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells[J]. Nature, 2013, 504(7480): 446-450. doi: 10.1038/nature12721 [6] MACIA L, TAN J, VIEIRA AT, et al. Metabolite-sensing receptors GPR43 and GPR109A facilitate dietary fibre-induced gut homeostasis through regulation of the inflammasome[J]. Nat Commun, 2015, 6: 6734. doi: 10.1038/ncomms7734 [7] MACHIELS KM, JOOSSENS J, SABIN O, et al. A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis[J]. Gut, 2014, 63(8): 1275-1283. doi: 10.1136/gutjnl-2013-304833 [8] TAKAISHI H, MATSUKI T, NAKAZAWA A, et al. Imbalance in intestinal microflora constitution could be involved in the pathogenesis of inflammatory bowel disease[J]. Int J Med Microbiol, 2008, 298(5/6): 463-472. http://pdfs.semanticscholar.org/b5ab/61c4449dfd4e8185eede697e5da3d08fac24.pdf [9] LOUIS P, DUNCAN SH, MCCRAE SI, et al. Restricted distribution of the butyrate kinase pathway among butyrate-producing bacteria from the human colon[J]. J Bacteriol, 2004, 186(7): 2099-2106. doi: 10.1128/JB.186.7.2099-2106.2004 [10] LASERNA-MENDIETA EJ, CLOONEY AG, CARRETERO-GOMEZ JF, et al. Determinants of reduced genetic capacity for butyrate synthesis by the gut microbiome in Crohn's disease and ulcerative colitis[J]. J Crohns Colitis, 2018, 12(2): 204-216. doi: 10.1093/ecco-jcc/jjx137 [11] BORTON MA, SABAG-DAIGLE A, WU JK, et al. Chemical and pathogen-induced inflammation disrupt the murine intestinal microbiome[J]. Microbiome, 2017, 5(1): 47. doi: 10.1186/s40168-017-0264-8 [12] SINGH N, GURAV A, SIVAPRAKASAM S, et al. Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis[J]. Immunity, 2014, 40(1): 128-139. doi: 10.1016/j.immuni.2013.12.007 [13] CHEN XQ, LYU XY, LIU SJ. Baitouweng decoction alleviates dextran sulfate sodium-induced ulcerative colitis by regulating intestinal microbiota and the IL-6/STAT3 signaling pathway[J]. J Ethnopharmacol, 2021, 265: 113357. doi: 10.1016/j.jep.2020.113357 [14] MIAO Z, CHEN L, FENG H, et al. Baitouweng decoction ameliorates ulcerative colitis in mice partially attributed to regulating Th17/Treg balance and restoring intestinal epithelial barrier[J]. Front Pharmacol, 2020, 11: 531117. http://www.researchgate.net/publication/348437836_Baitouweng_Decoction_Ameliorates_Ulcerative_Colitis_in_Mice_Partially_Attributed_to_Regulating_Th17Treg_Balance_and_Restoring_Intestinal_Epithelial_Barrier [15] HUA YL, JIA YQ, ZHANG XS, et al. Baitouweng Tang ameliorates DSS-induced ulcerative colitis through the regulation of the gut microbiota and bile acids via pathways involving FXR and TGR5[J]. Biomed Pharmacother, 2021, 137: 111320. doi: 10.1016/j.biopha.2021.111320 -
 下载:
下载:



 点击查看大图
点击查看大图

计量
- 文章访问数: 213
- HTML全文浏览量: 135
- PDF下载量: 72
- 被引次数: 0
